‘Sunburned’ Seaweed - an increasing phenomenon on the Atlantic coast of Nova Scotia?
by Debbie MacKenzie April 7, 2002
A pattern of dying seaweed, that at first glance looks like a case of ‘sunburn,’ has lately become evident in Nova Scotian seaweeds. This development is consistent with changes that would be predicted to occur "IF" (as the theme of this website contends) a gradual lowering of nutrient availability in the open ocean were to occur…
Brown seaweed that is cast adrift and dries out above the high tide mark soon becomes dried and blackened, and anyone who walks the shoreline is familiar with this fact…but the recent (springtime) appearance of dry, red-black scorched tissue on long-lived seaweeds that are still alive, still attached to the rocks, seems to present another sign of the general deterioration (perhaps "starvation?") of the marine ecosystem…this change is not a part of the ‘normal’ annual life cycle of these marine plants, but a signal that they are experiencing an intolerable level of some type of stress..
What is the root cause of this picture?
August 4, 2001. - My attention was first drawn to the gross development of deep red patches in this bed of yellow-green seaweed at East Dover, N.S., last summer (photo at left). Email inquiries to seaweed specialists revealed their agreed opinion that the red tissue was dead and that it was a normal occurrence which they have observed when this common seaweed, Ascophyllum nodosum, is exposed to unusually intense heat, usually during mid-day low tides in summer. Although there was no unusually hot weather in this part of Nova Scotia during the summer of 2001 that might have triggered the appearance of the red spots, I actually had photographed the reddened seaweed on a sunny day with low tide at mid-day…so I left it at that. The extreme appearance of the red spots did not last very long, as checking the same site several days later revealed that the ‘spots’ had faded to a lighter greyish color, offering much less contrast with the yellow background. After that time, the greyish spots disappeared, presumably as the dead tissue was sloughed off, leaving the same yellowed plants with yellowed tips - just cropped several inches shorter than before the episode of ‘sunburn’ or "heat" damage.
Seven months later, March 29, 2002, and again I see "heat" damage in Ascophyllum nodosum.
During the last week of March, we had a series of mid-day low tides. The weather was average for early spring in Nova Scotia (see long term temperature data graph included with references at bottom of page). Temperatures fluctuated between a low of a few degrees below zero and a high of about 8 degrees Celsius; there were some sunny periods, almost constant wind, occasional snow flurries and occasional rain. By no stretch of the imagination was there a "hot" day during this week (or indeed, during the 6 months that preceded this week), yet it appears that Ascophyllum plants have recently sustained new "heat" damage here, possibly during this latest series of mid-day low tides. Some factor besides "heat" is clearly affecting the seaweed.
The following pictures were taken in several sheltered locations, at Prospect, East Dover and Peggy's Cove, between March 29 and April 5, 2002. The weather has remained cold and windy.
March 29, Prospect, Nova Scotia.
Close view of damaged Ascophyllum at Prospect, April 5.
Severely damaged specimen at Prospect.
Perspective from sheltered inlet at Prospect - the innermost plants, showing yellowing, are those now affected by the "heat" damage.
Damaged Ascophyllum near Peggy's Cove.
Some longer strands of seaweed are also affected at East Dover.
Damaged Ascophyllum growing high on a sheltered rocky reef at East Dover - April 5.
Close view of damage on the East Dover reef.
The rocky reef surveyed at East Dover. At this distance the red tinge is barely visible in the most yellowed Ascophyllum to the right of the center of the reef. (And this photo has obviously been taken during late winter/early spring - note the lack of foilage on deciduous trees in the background).
Approaching high tide at the East Dover reef - note the still water in the area where the most severely yellowed and "heat" damaged seaweed grows.
The pattern is predictable.
The vast majority of Ascophyllum living on the shoreline is NOT exhibiting the red-black scorched look - this damage is limited to plants living at two extremes: (1) the upper vertical limit of its range and (2) in the most sheltered locations, where plants experience the lowest degrees of water motion. Both of these variables contribute to minimizing the availability of nutrients in the seawater to the seaweeds. Plants living at the upper vertical limit of their range have the shortest immersion (feeding) times. And plants living in relatively still water have less opportunity to absorb nutrients from seawater since they can deplete the water layer that is immediately adjacent to their fronds.
Besides immersion time and degree of water motion, a third variable, the actual nutrient concentration of the seawater, also obviously plays a key role in determining the extent of the range that is habitable by these plants. This seems most likely to be the variable that has recently changed for the worse.
Ascophyllum is a long lived perennial plant, with individuals capable of surviving for up to six decades or more. This seaweed once successfully established and sustained itself in these sheltered inlets, and obviously received adequate nutrients from the tides to support its growth, but it is now breaking down there. Immersion times and water flow patterns in these areas have not undergone any recent changes (the tidal rhythm remains predictably constant), which leaves a decline in nutrient content of the seawater as the most plausible explanation for the recent deterioration.
How could a scarcity of nutrients lead to "heat" damage?
Malnutrition is well known to lower the natural resistance or defenses of organisms, both plants and animals, to stressors that naturally occur in their environments. For plants, malnutrition raises susceptibility to thermal stress, damaging effects of light, infectious diseases and parasites. For seaweeds, especially those situated at the higher positions in their habitat, dessication is a significant environmental stressor. In fact, dessication tolerance is believed to be the limiting factor which determines the upper limit of where Ascophyllum can live in the intertidal zone (Lobban and Harrison, 1994). Often a sharply delineated horizontal line defines the upper limit of Ascophyllum growth as a result. Scientists have determined that the loss of 70% of tissue water content predictably results in cell death in this seaweed (DFO, 1997).
It has also been determined that the thermal tolerance for Ascophyllum is 26 degrees Celsius (DFO, 1997) - but since temperatures nowhere near this limit could have affected these plants for many months, it is far more likely that the dessication threshold is the one that was crossed. A general weakening of the plant due to poor nutrition would predictably lower all natural defenses, including natural resistance against extreme dessication. (The year-round yellow ‘bleached’ coloring of these plants also reflects their poor condition as normal (olive green) pigments are lacking.)
Might the ‘sunburn’ or "heat" damage pattern actually be a symptom of the damaging effects of ultraviolet radiation?
No. This is really not plausible for several reasons: (1) plants of the same species with equivalent exposures to air and light (but higher nutrient availability due to spending their (equally short) immersion times in moving water) are not showing the same pattern of damage. (2) If UV light were to increasingly become a stressor for plants, the earliest manifestations of ill effects would be expected to be seen during the time of year when sunlight is most intense (summer). Marked damage occurring during late winter and early spring due to UV radiation would simply not be expected…or at least not before major damage had been inflicted in summertime.
Consider dessication: the loss of 70% of water content from tissue is lethal to Ascophyllum nodosum (DFO, 1997).
Which factors intensify dessication?
(1) Heat significantly accelerates drying, which is probably partly why I observed "heat" damage last summer (even though the known thermal threshold for the plant had not been crossed ). But "heat" in March cannot be the factor that has intensified dessication.
(2) Wind can also rapidly accelerate drying. Could wind be the factor that has caused the extreme dessication stress for the seaweed at this time? Our recent weather has included quite a few windy days, although nothing outside of what would be considered normal for this time of year.
(3) Relative humidity of the air also affects the rate of drying.
Dessication stress experienced by seaweed at this time of year might be as severe (or even more so) than that experienced during the warmer summer months for two reasons: (1) Winds are significantly more intense during winter and spring and (2) relative humidity levels are significantly lower during these months as compared to the summer months.
Dessication stress is determined by the combination of temperature, wind speed and relative humidity, and can be intensified by extreme values of any of these three variables. Dessication resistance in seaweeds is a function of emersion (air exposure) time and the health and strength of the plant. If climate-related environmental factors remain constant but the plant becomes significantly weakened, emersion time will need to be decreased to compensate. Since the plants cannot move, the most intensely stressed ones will die. This explains why the highest positioned Ascophyllum plants are dying in the sheltered inlets, and why the immediate cause of death appears to be ‘sunburn’ or "heat" damage although the primary underlying cause is nutrient starvation.
What is the significance of this localized deterioration in Ascophyllum?
In late March-early April 2002, observations were made of similar habitat type (high intertidal zones of clean, sheltered inlets) at various points along approximately 30 kilometres of shoreline (from Peggy’s Cove to Prospect), and the pattern of "heat" damage in Ascophyllum was consistently found. Presumably a much wider area than this has been similarly affected.
A few relevant points:
- Seaweed communities have been considered to be useful indicators of nutrient enrichment of coastal waters. Their sessile, long lived nature provides several advantages as their condition integrates fluctuations in nutrient supply over long terms much better than shorter lived species such as phytoplankton. These same qualities also make seaweeds excellent biomonitors for nutrient impoverishment of seawater. However there has been no index of suspicion on this point, and research efforts to date have been largely focused on seaweeds in grossly polluted estuaries (Schramm and Nienhuis, 1996).
- The decline (or disappearance) of similar brown seaweeds, fucoids, has been noted in other parts of the world. There is good evidence that severe eutrophication can lead to the disappearance of some of these species (one well known example is their practical elimination from the Baltic Sea (Schramm and Nienhuis, 1996)), less dramatic declining trends have been noted in other areas…and in less obviously nutrient enriched areas…might nutrient depletion also be an unsuspected factor contributing to their decline in some areas?
- There appears to have been no index of suspicion on the part of scientists that changes might be induced in intertidal communities by a declining availability of nutrients or decreased nutrient cycling in the ocean. And practically no long term studies have been undertaken in unpolluted intertidal zones - in Atlantic Canada, for instance, monitoring of seaweeds is in its infancy (Bates et al, 2001, Chopin, undated). Early reports of recent baseline monitoring here have noted "symptoms of stress" in some seaweeds in the Bay of Fundy - but the interpretation has initially been posited as being related to increases in terrestrial-sourced organic input (Bates et al, 2001). Dramatic changes in intertidal communities have been recorded in well documented cases elsewhere in the world, but the changes are consistently discussed in relation to either nutrient enrichment or global warming (e.g. Barry et al, 1995, Schramm and Nienhuis, 1996)
- The starvation of seaweeds in small sheltered inlets that are flushed tidally by the (unpolluted) water of the North West Atlantic ocean may offer a very important clue as to the exact pathology of ‘what is wrong’ with our failing marine life. The assumption of a steady availability of nutrients and ‘constant productivity’ of this ocean may no longer be justified. (And this one is not a story about the effects of global warming...)
Nutrient depletion of the seawater - how might this develop?
There seem to be two basic possibilities:
(1) It is a natural cycle.
The availability of dissolved nutrients in seawater in temperate zones is well known to fluctuate quite widely on an annual basis. This is especially true of the surface layer, the part of the water column that sustains the seaweeds in question. As the surface water warms in summer it is well known to become relatively stripped of dissolved nutrients, but the low temperatures and higher winds in winter induce mixing of the water column, which raises surface water nutrient content to its highest annual concentration. Therefore the observed damage sustained by the seaweed appears, paradoxically, to have occurred during the season of its highest nutrient availability. The condition of these plants, however, does not reflect instantaneous nutrient concentration. The health of the longer lived seaweeds is more a reflection of an integrated average of long term nutrient supply (Chopin, undated, Shubert, 1984). These perennial seaweeds normally store nutrients in their tissues, reserves that are absorbed during times of higher availability and used for growth in seasons of lower availability (Lobban and Harrison, 1994). Inadequate nutrient availability on an annual basis will weaken these plants to the point that occurrence of intense natural stressors, such as dessication, will cause death regardless of the season, or the immediate concentration of nutrients in the seawater. Dying on the rock, especially during late winter/early spring in Nova Scotia, is definitely not a part of the natural life cycle of Ascophyllum nodosum.
(2) A second possibility is that a lowering of overall nutrient availability in the ocean has occurred because too many nutrients have been removed…and possibly removed in the form of fish. Systemic nutrient depletion could be a gradual, long term, cumulative effect of fishing removals, if effective replacement has not occurred. This is not an unknown concept, and the possibility has been described in standard texts, as in this excerpt from "Biological Oceanographic Processes" (Parsons et al, 1984):
"The amount of fish taken out of any ecosystem must be proportional to the amount of production within that ecosystem. Since nitrogen is generally regarded as the rate-limiting element in many (but not all) ocean ecosystems, it is appropriate to consider first how much new nitrogen enters an ecosystem each year. The alternate source of this element is recycled nitrogen. However, recycled nitrogen is largely retained within the system so that removal of nitrogen from such a closed system in the form of fish results in an ultimate decimation of the nitrogen reserves of the ecosystem." (Parsons et al, 1984)
Production resulting from vertical mixing has therefore been considered "new" production…although looking at the wider picture reveals that these nutrients have just been "recycled" via a longer route…so an ecologically significant difference may not exist in the system between what we perceive as two different types of production ("new" and "recycled"). Sea life may simply have evolved to base all production on "recycled nutrients," just using a variety of recycling pathways.
Scientific thinking in fisheries management has traditionally been that "surplus production" exists (due to the continuous supply of "new" nitrogen) and that fish removal at an appropriate level will therefore not result in nutrient depletion or serious negative impact on the system. However, the marine ecosystem appears to be in serious disequilibrium today…and maybe basic views like this one now need to be reassessed.
All of the fish removed by humans would have been "naturally recycled" had the fishing not occurred. We reassure ourselves however, that we have poured tons of nutrients into the sea as "replacements." The forms of replacement nutrients however, (mainly sewage and fertilizer), do not appear to be effectively cycled throughout the web that is subjected to fishing. Therefore the distinct possibility exists that a reduction of the nutrient inventory of the marine food web has resulted directly from centuries of fishing (…and this has been argued in great detail elsewhere on this website).
Regardless of theoretical arguments that might be made against the hypothetical development of nutrient depletion in the open ocean, the cold observation remains that Ascophyllum nodosum is dying back in springtime in clean sheltered inlets along the open North Atlantic coast. This is not "normal." And yet, the decline of Ascophyllum is not particularly surprising when viewed alongside other recent changes in coastal marine life, changes that seem to point in the same direction. For instance, look at the change that has occurred in the population of barnacles in this area. The loss (contraction of vertical range) of these common intertidal organisms is very hard to account for by using other models of how the marine ecosystem works.
And the "starving ocean theory" clearly now deserves serious attention.
References
Barry, J. P., C. H. Baxter, R. D. Sagarin and S. E. Gilman. 1995. Climate-Related, Long-Term Faunal Changes in a California Rocky Intertidal Community. Science 267: 672-675.
Lobban, Christopher S. and Paul J. Harrison. 1994. Seaweed Ecology and Physiology. Cambridge University Press.
Parsons, T. R., M. Takahashi and B. Hargrave. 1994. Biological Oceanographic Processes (third edition). Pergamon Press.
Schramm , W. and P. H. Nienhuis (eds) 1996. Marine Benthic Vegetation, Recent Changes and the Effects of Eutrophication. Springer, Ecological Studies 123. (470pp)
Shubert, L. Elliot (ed). 1984. Algae as Ecological Indicators. London: Academic Press, Inc.
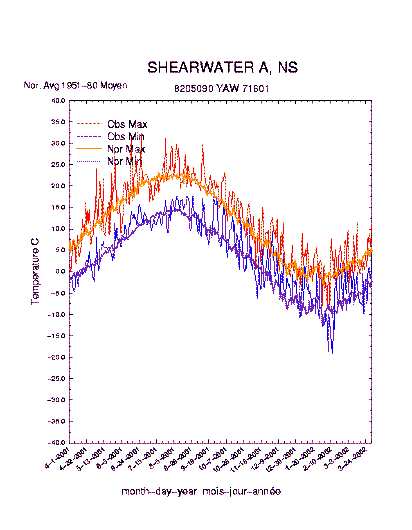
At left: Recent temperatures compared to long term mean. Graph from Environment Canada.
For more detail on changing trends in seaweeds, see seaweed article posted on this site last summer, and also seaweed update posted late December, 2001.

 August 4, 2001. - My attention was first drawn to the gross development of deep red patches in this bed of yellow-green seaweed at East Dover, N.S., last summer (photo at left). Email inquiries to seaweed specialists revealed their agreed opinion that the red tissue was dead and that it was a normal occurrence which they have observed when this common seaweed, Ascophyllum nodosum, is exposed to unusually intense heat, usually during mid-day low tides in summer. Although there was no unusually hot weather in this part of Nova Scotia during the summer of 2001 that might have triggered the appearance of the red spots, I actually had photographed the reddened seaweed on a sunny day with low tide at mid-day…so I left it at that. The extreme appearance of the red spots did not last very long, as checking the same site several days later revealed that the ‘spots’ had faded to a lighter greyish color, offering much less contrast with the yellow background. After that time, the greyish spots disappeared, presumably as the dead tissue was sloughed off, leaving the same yellowed plants with yellowed tips - just cropped several inches shorter than before the episode of ‘sunburn’ or "heat" damage.
August 4, 2001. - My attention was first drawn to the gross development of deep red patches in this bed of yellow-green seaweed at East Dover, N.S., last summer (photo at left). Email inquiries to seaweed specialists revealed their agreed opinion that the red tissue was dead and that it was a normal occurrence which they have observed when this common seaweed, Ascophyllum nodosum, is exposed to unusually intense heat, usually during mid-day low tides in summer. Although there was no unusually hot weather in this part of Nova Scotia during the summer of 2001 that might have triggered the appearance of the red spots, I actually had photographed the reddened seaweed on a sunny day with low tide at mid-day…so I left it at that. The extreme appearance of the red spots did not last very long, as checking the same site several days later revealed that the ‘spots’ had faded to a lighter greyish color, offering much less contrast with the yellow background. After that time, the greyish spots disappeared, presumably as the dead tissue was sloughed off, leaving the same yellowed plants with yellowed tips - just cropped several inches shorter than before the episode of ‘sunburn’ or "heat" damage.