
|
|
CONCLUSIONS RE: MASS CORAL BLEACHING - IT’S TIME TO “THINK OUTSIDE THE BOXES” - (and consider this one: BY UNDERMINING THE “BIOLOGICAL PUMP,” FISHING IS HAVING A DELETERIOUS EFFECT ON THE ATMOSPHERE) Reef corals are disappearing at an alarming rate; this has taken us by surprise; our scientific understanding, and even basic assessment of the state of the world’s reefs is inadequate. (Reefcheck) “Human populations dependence on reef services is very unfortunate, that we’ve come to have such a dependence on the sea and our demands are now so great, just as the well is showing clear signs of running dry.” - Rafe Pomerance “Could reef scientists possibly make rational arguments for considering the entire ecosystem as endangered...including commercially valuable fish and shellfish which play a role in nutrient distribution and recycling, etc.?” (question posed to the “coral-list” during a discussion on the proposed ESA listing for one coral species) Those are exactly the “rational arguments” offered by this report. The “entire ecosystem” clearly is endangered...AND commercially valuable fish play a critical role in “nutrient distribution and recycling.” Coral reefs have been suggested as microcosms for the larger marine ecosystem - and, given their nearly self-contained nature, this is a reasonable concept - the alarming thing now is that these microcosms are showing serious damage at the very “foundation” of the system. Unexpectedly, the corals are now dying - does this parallel the crumbling of the very foundation of the larger marine ecosystem? In all likelihood, it does. It’s time for us to bite the bullet, however difficult, and take an “ecosystem approach” to “managing” the ocean. All the subtleties of the inter-relationships between marine organisms we can never hope to appreciate - but we do know that the inter-dependencies exist, so we must now acknowledge that the exploitation of any member of the “web” has an impact on the system as a whole. Fishing ultimately decreases “everything” in the sea. The single species approach will simply never succeed in “rebuilding” and “conserving” fish stocks. -- Converging lines of evidence seem to support the hypothesis that corals expiring in “mass bleaching events” are dying because they have been severely weakened by undernutrition. The physical changes, including weight loss, decreased immunity, low lipid stores and the lowering of thermal and irradiance tolerances, are identical to the pathophysiology of simple food starvation in corals. This needs to be investigated, and designing appropriate experiments should be a relatively simple matter. -- There is also good reason to suspect that fishing is an independent risk factor in coral bleaching. This hypothesis also needs to be promptly investigated. (It’s time to overcome our natural reluctance to ask this question - denial can be deadly.) -- The critical role of fish in nutrient recycling in coral reef systems, provides the theoretical basis to propose fishing as the ultimate cause of starvation in corals. (Ditto for the ocean system as a whole.) -- Much more work, especially including original approaches, needs to be done regarding the “marine nutrient cycle.” One very significant aspect of “nutrients” in the sea is their physical form - solid vs. liquid - limiting the “nutrient” concept to the dissolved and particulate (liquid) forms, constrains our thinking and hampers our understanding of the system. “Fish” are obviously “nutrients” (why else would we catch and eat them?) -- And we are forced to work with incomplete information. Time-series biological data appears to be very scarce in tropical marine science. Single measurements are just that, and must not be assumed to define constant properties. One example is the “productivity” of coral reefs - gms carbon fixed/area/time - while this may be difficult to calculate, once it’s done we still have only one measurement. In an obviously changing system, it’s illogical to assume that a figure like “primary productivity” will be a constant. Another example is the “thermal thresholds” of corals - taken from their behavior during recent bleaching events, there is no reason to assume that these absolute thermal tolerances are inherent characteristics of the corals. Rather it is more likely that “primary productivity” and “thermal thresholds” are both just single points on two more as-yet-undescribed downward sloping graphs - that’s really where the evidence points, since fishery yields and heat tolerances of corals both were evidently higher in the past. THINKING “OUTSIDE THE BOX” - WHERE ARE THE “BOXES?” In essentially all human endeavors, we have a tendency to internalize and accept familiar ways of seeing and thinking about things. Eventually we somehow come to believe that these ways of thinking, since they are so widely accepted, must be based on underlying “truths.” That’s when most of us stop questioning them. It might be a classification system that once appeared to make sense; often however, it’s the unconscious human bias that’s hidden in our language, in the labels that we choose to apply to things. These are examples of human-created “boxes,” a few of the ones that we need to try to “think outside of.” 1. “Predator” “Prey” “Herbivore” “Carnivore” “Omnivore” “Scavenger” “Primary producer” “Nutrient” “Food Chain” “Commercially significant species”.......these are a few of the “boxes” used in marine biology. The labels “predator” and “prey” are interesting. These define roles played by organisms in natural ecosystems. “Predator” defines the fish by who he kills and eats. “Prey” defines a fish by who he feeds, who kills and consumes him. So we define them by what they eat (who feeds them) and by what eats them (who they feed). So each creature therefore warrants two descriptive labels. Descriptions of food web dynamics will include this typical example: HERBIVORES are PREY for CARNIVORES. There are two words there for the smaller fish, “herbivore” (what feeds him) and “prey” (who he feeds). But there is only one word for the “carnivore” (who feeds him). Lacking from our vocabulary is the word to describe “who” the carnivore naturally “feeds.” We know that the food web is an endless cycle, energy constantly flows through while scarce nutrients are carefully recycled. We understand also how it is that the top carnivore provides nutrients to organisms “lower” in the web. (“Lower” and “higher” are actually just two more human-bias “boxes.”) We know that fish excrete ammonia and urea which are used to fertilize phytoplankton, we also know that fish feces are consumed directly by smaller species, as are millions of excess eggs and larvae that are produced by the bigger fish, and that, even if they have not “killed” it, smaller species are very quick to consume carrion remaining from dead “top predators.” When the capelin consumes the eggs of the cod...have they both stepped outside of their assigned “boxes?” Yet we lack a word to capture these natural functions of the carnivore, the completion of the “downside” of the food web -- the label that we apply to carnivorous fish to describe “who he feeds” is “COMMERCIALLY SIGNIFICANT SPECIES.” In other words, he feeds “us.” Of course he does feed us, but it’s a new development in his history, and it’s not “natural,” in fact it breaks the long-established marine nutrient cycle, and our language deficit reflects our inability to acknowledge this fact. The carnivore’s natural destiny or obligation (in the sense of duty that we assign to those we label “prey”), is not to feed humans, but to feed an assortment of smaller marine organisms. What is missing from our language is a word to describe the other natural “feeding” function of the carnivorous fish, a word that conveys the idea of his role as “food processor and distributor for smaller organisms.”  “Predator/prey” is just one example of “boxes” built by human thinking which necessarily
contain human biases. We seem to leave the “upper” end open beyond the “top predator” -
in the food web diagrams he generally appears to be something of a dead end - is that
because, at some level, we don’t want to acknowledge that this big fish “owes” something
to the system as a whole? And we like to think of ourselves as just another “top predator”
in the marine ecosystem. If we acknowledged the obligation of “top predators” to properly
complete the downside of the “food web,” we might be forced to take a hard look at
exactly what it is that WE are “giving back” to the system. And this might become
uncomfortable...but denial is a great defense...for a short while... Undoubtedly, one factor
determining the initial creation of the “boxes” of “predator,” “prey,” and “upper and lower
levels” of organisms - was human visual acuity. Originally it may just have reflected what
was most easy for us to see with our eyes. We actually know a lot more about the
ecosystem now, but are still using the same old mental “boxes.”
“Predator/prey” is just one example of “boxes” built by human thinking which necessarily
contain human biases. We seem to leave the “upper” end open beyond the “top predator” -
in the food web diagrams he generally appears to be something of a dead end - is that
because, at some level, we don’t want to acknowledge that this big fish “owes” something
to the system as a whole? And we like to think of ourselves as just another “top predator”
in the marine ecosystem. If we acknowledged the obligation of “top predators” to properly
complete the downside of the “food web,” we might be forced to take a hard look at
exactly what it is that WE are “giving back” to the system. And this might become
uncomfortable...but denial is a great defense...for a short while... Undoubtedly, one factor
determining the initial creation of the “boxes” of “predator,” “prey,” and “upper and lower
levels” of organisms - was human visual acuity. Originally it may just have reflected what
was most easy for us to see with our eyes. We actually know a lot more about the
ecosystem now, but are still using the same old mental “boxes.”
2. “The food chain” “the food web” “the BASE of the food chain/web” Phytoplankton, the “primary producers” who accomplish the critical role of harnessing energy from sunlight and carbon fixation, are commonly described as the “base” of the “food web.” Their role is a crucial one to be sure, but calling them the “base” implies that they somehow stand alone as a foundation that is capable of supporting everything above it. In the sense of “energy capture” this is true, but regarding “nutrients” it is not. Maybe it would be helpful to isolate the concepts into the “energy web” and the “nutrient web” instead of lumping them together as the “food web?” It adds to the confusion. The typical food web diagram shows top predators as dead ends - that’s not true even if the web is reflecting energy alone, since it’s well known that fish assimilate only a minor fraction of the energy and nutrients that they consume - a significant fraction of energy and nutrient content both are redirected “downward” in the web - and these arrows are most often missing from the diagrams. A diagram of the “nutrient web,” especially one focussing on fixed nitrogen, would show a complex pattern of transfers between organisms big and small, all playing their roles and taking their turns, but nothing - certainly not phytoplankton - would stand alone as the “base,” something capable of “supporting” the system. Therefore the “base of the food web” is another human-built box for the phytoplankton. Also, might our labelling them as “primary producers” imply that somehow, on their own, phytoplankton have the ability to “produce” fish? 3. “Overfishing” and “sustainable fishing” These are two of our verbal “boxes” that have been giving us a lot of trouble lately. We need these to perpetuate the myth that removing fish from the sea (without appropriate nutrient replacement), can be a sustainable human practise over the long term. We insist on thinking that “overfishing” is an inherently different activity than “sustainable fishing.” “Overfishing,” we realize will result in the depletion/disappearance of the target fish. We’ve seen many blatant examples up to this point, so we do believe that “overfishing” can be destructive. But we persist in thinking that another type of fishing “must” exist that causes no harm - and we’ve labelled that box “sustainable fishing.” It’s the goal that continually eludes us however, we can’t quite make it, and we find ourselves having to continually lower the hoop. It’s become obvious that levels of exploitation that appeared “sustainable” in the past, simply are not so now. Was the “sustainability” of earlier fishing an illusion, produced when a relatively low level of fishing was applied to a very rich and therefore resilient, resource? We do not have a clear enough understanding of all the effects of fishing - and still for the most part imagine fishing affecting only the target species (or at most the target plus it’s usual immediate predator) - however “fishing” clearly has repercussions throughout the whole ecosystem, and all “non-targetted” or “unexploited” species ultimately feel the effects. “Overfishing” and “sustainable fishing” represent an arbitrary division that we have chosen to place in the “fishing” box - for our own purposes, it carries much less significance for the larger ecosystem. The balance of individual species will be affected more or less by fishing effort targeted at one or the other, but the ecosystem as a whole will react and adjust to the overall cumulative effects of “fishing,” regardless of how we choose to think about it. Mass coral bleaching provides a prime, “easy-to-see” example of how marine ecosystems as a whole react to our relentless fishing - the whole thing becomes undermined and starts to fall apart, bleached corals are showing us how “unexploited” marine species ultimately suffer the terminal effects of nutrient depletion initially caused by “fishing.” They are dying from starvation in the oceanic “microcosm.” (Actually, tiny first and second order “consumers” in temperate ecosystems are also disappearing - zooplankton are in decline - it’s just that those examples do not leave behind the glaring white skeletons that the corals do, so the alarm bells are not sounding as loudly there.) 4. “Nutrient overload” “eutrophication” “harmful effects of nutrients added to coral reef systems” “global nitrogen glut” While excessive amounts of nutrient-laden liquid runoff does cause damage to the environment, this constant theme distracts us from the possibility that coral systems might experience a nutrient deficit. The feeling almost comes across that corals are somehow “allergic” to nutrients - occasionally, (although it’s sloppy reporting to be sure), media sources will actually print statements like “corals need clean, nutrient-free water.” Has our concern about the damaging effects of liquid coastal nutrient inputs caused us to miss the problem of nutrient deficiency that is developing in other marine systems? Corals will predictably die of starvation in water that is literally “nutrient-free.” Have our thoughts on corals and nutrients all been placed into a “box” labelled “Warning! - avoid harming coral reefs by adding nutrients?” And we seem to have placed “nutrients” and “fish” into two separate mental boxes. Is this because it suits our purposes to think of the pool of “nutrients” that supplies the food web as being something distinctly separate from the actual fish? Well, fish ARE nutrients, it’s true everywhere, but is particularly well illustrated in the tropical systems. It is more obvious there that removing a significant bulk of the fish lowers the nutrient content of the ecosystem as a whole...the clear tropical water is merely the best example to destroy the myth of the “vast pool of available nutrients” in the sea. 5. Functions of ecosystems - in which ways can ecosystems be thought of as “organisms?” Like individual organisms, natural ecosystems have built in homeostatic, or stabilizing, mechanisms - processes that “kick in” to maintain relative stability when conditions shift towards extremes. There is concern today about the “global nitrogen overload.” Since the development of industrial methods to fix atmospheric nitrogen for fertilizer, the gross global amount of fixed nitrogen has increased markedly, according to the statistics. Greatly increased amounts of this fertilizer have been flushed into the sea - hence the conclusion (myth) that the sea has an “oversupply of nutrients.” It has even been calculated that this nitrogen input far exceeds the nitrogen output from the sea via fishing removals. But what has really been happening with all that nitrogen? Living systems can only tolerate fixed nitrogen “fertilizer” up to a certain concentration - above that point it becomes counterproductive, even harmful to the same organisms that would normally benefit from it. Terrestrial ecosystems have a method to rid themselves of excess fixed nitrogen - flush it downstream. Rainfall washes the excess liquid nitrogen from the land into streams and rivers where it will ultimately be carried to the sea. (“Aerial deposition” of nitrates has the same end result.) When faced with a too-high concentration of liquid fixed nitrogen, the aquatic system does not have the option of tossing it back to the land, so other methods have been built in to deal with it - to effectively “get rid of it” and minimize the damage. Nitrates carried to the sea via freshwater are dealt with in the following ways - (1) The initial salinity gradient causes precipitation/flocculation of a certain amount, which then sinks to the sediment, (2) Phytoplankton bloom more profusely than normally, consuming a large portion of the nitrogen, dead plankton then sink to the bottom where bacterial decomposition uses up the oxygen in the water. (3) As the water becomes increasingly hypoxic, the activity of the anaerobic denitrifying bacteria increases, and they busily convert the fixed nitrogen back to N2, which is returned to the atmosphere as a gas. This is similar to the working of a septic system, and these natural processes work to rid the seawater of excess nitrogen by effectively adding it to the bottom sediment or returning it to the atmosphere. High coastal liquid nutrient input therefore does not result in a “well nourished” marine ecosystem, nor does it contribute to maintaining “healthy fish stocks.” The ocean “doesn’t like it”...so the ocean does what it can to “get rid of it.” That’s what happens to liquid nutrients - solid nutrients, on the other hand, receive an entirely different treatment in the sea - large solids are most acceptable, a dead whale for example is a real boon to the community of scavenging organisms nearby. Constantly nibbled at by many mouths until it’s gone, bacterial colonization of the larger surface never gets very well established. Therefore the bulk of the nutrients in the dead whale enters the living food web to be recycled - only a minor fraction of the nitrogen would wind up in the bottom sediment or back to the atmosphere, and the process of consuming/breaking down the carcass should not particularly provoke the development of hypoxia in the area. Thus there is a “world of difference” between adding nutrients to the sea in solid or liquid form; and this is why this author argues that we should “feed the fish” with solid food - our food scraps, slaughterhouse waste, unwanted dead livestock (good whale substitutes?)...whatever we have that contains protein that living sea creatures could consume. Regarding the “starving” coral reef ecosystems, unsightly as we might find it, the best way to help restore their health might be to place livestock carcasses around their edges (or at least it’s worth a try on one “experimental” reef - test whether or not corals will eventually become more bleaching resistant if the ecosystem is fed - it might take a while for the effect to be seen, but it’s got to be better than standing by and just watching the bleaching problem get progressively worse. And it will not cause any harm, dead animals in the sea do not cause “eutrophication;” they are a very natural part of the normal picture.) Those interested in rebuilding marine fish stocks (including enhancing the health of sick corals) would do well to take a page out of the book of some Pacific salmon researchers. Realizing that the stream ecosystems are feeling nutrient loss (“starving”) since the fish are now virtually absent, they have been taking the (successful) step of tossing the carcasses of dead hatchery fish into the streams - for the purpose of nourishing the 100-plus species that normally benefited from the natural death of fish in the area. Applying liquid fertilizer in this scenario would obviously be inappropriate; rather than nourishing the juveniles and others, it would stimulate an unnaturally high phyto-bloom and trigger the “get rid of it” compensatory response already described for cases of “eutrophication.” So the sea has a built in strategy to counteract situations wherein dissolved nutrient levels become too high. What happens if they drop too low? Are there compensatory mechanisms built in for that possibility? It seems that there might be. The class of organisms known as “blue-green algae” or “cyanobacteria” are unique in the sea in that they have the ability to make use of atmospheric nitrogen. Called “nitrogen fixers,” their accumulated work over the millions of years of the history of life in the sea, is credited with amassing the wealth that was once there. “Nitrogen fixers” find themselves at a competitive advantage over other types of algae when the nitrogen concentration becomes very low in the water. Their trump card is that fact that they can “make their own fertilizer.” Blue-green algae are a significant feature of coral reef ecosystems, commonly forming what is known as “turf algae.” With the recent decline in nutrient levels, this may possibly be more prevalent than it once was (a possible compensatory mechanism for fishing removals? Has the growth of the blue green algae increased on the exploited coral reefs? Maybe. If so, it’s the only way that the ecosystem could attempt to stabilize its biomass/nitrogen content once subjected to depletion by human fishing.) Interestingly, elsewhere in the ocean the occurrence of certain forms of blue green algae is definitely increasing. Blooming in calm, warm, nutrient-stripped offshore waters, some of these are the causative agents of the toxic “red tides,” the “harmful algae blooms” that are becoming an increasing problem. All “toxic” algae blooms cannot be grouped together, and there are certainly those that bloom in nearshore nutrient-loaded waters, for example Pfeisteria. But there are a range of toxic dinoflagellates, that are capable of N-fixation, that are blooming when water, often offshore, becomes nutrient-stripped - as in warm, doldrum conditions - and then drifts towards shore, poisoning fish, shellfish and sometimes their human consumers. It is a problem with an increasing frequency worldwide. These organisms produce some nasty toxins, small doses are capable of killing fish and mammals. “Why” might they do this? It has been demonstrated that at least one of these organisms, Alexandrium fundyense, produces a much more concentrated toxin when grown in conditions of even lower nutrients (phosphate stressed). These blue-green generated toxins must have been designed to do what they do - i.e. kill fish and mammals. What purpose would be served for the ecosystem by killing these organisms? Well, it may be plausible that it is a systemic response to the lack of dissolved nutrients. Other phytoplankton cannot grow in the “too-clean” water, so the blue-green algae have very little competition for the light and the nutrients that are there. Capable of “fixing” the nitrogen shortage, but not the phosphate one, does it perhaps serve a corrective purpose for the system to kill some animals at this point, thereby releasing at least a portion of their nutrients into the water in dissolved form? A fish killed by the toxins might be more prone to bacterial breakdown, with release of nutrients into the water, than an ‘ordinary’ dead fish...since the toxin has probably killed many of the fish’s natural consumers as well. This result would help to “correct” the severe dissolved-nutrient depletion that triggered the toxin production in the first place....Just thinking...just wondering if this part of the increase in “harmful algae blooms,” the growth of toxic blue-greens in nutrient-stripped waters, might represent one of the checks and balances that were incorporated into the complex, persistent marine ecosystem on this planet - is the increasing level of fish-toxins a compensatory response to increasingly nutrient-depleted seawater? (Periods of calm, warm weather are not new or increasing, overall seawater nutrient levels offshore, however, are in all likelihood decreasing.) NOW, FINALLY, “THE REAL CONCLUSION” -- THE MOST “OUTRAGEOUS” OF THESE CONCLUSIONS, IF YOU HAVE READ THE EARLIER ESSAYS ON THIS WEBSITE, YOU KNOW THAT IT ALWAYS COMES DOWN TO THIS ONE....(step outside, wayyyy outside the “boxes”...) “THE RISE IN ATMOSPHERIC CARBON DIOXIDE LEVELS THAT IS CURRENTLY OCCURRING ON THE PLANET IS, TO A SIGNIFICANT EXTENT, A DIRECT CONSEQUENCE OF HUMAN REMOVAL OF LIFE FROM THE SEA” It is certainly not an original insight that decreasing growth of corals in the sea will predictably aggravate the accumulation of CO2 in the atmosphere. But it is true. “Climate protection” is sometimes listed among the “environmental services” provided by coral reefs: “Marine photosynthesizing and shell-forming organisms tie up carbon dioxide that would otherwise intensify global warming.” (WRI) “An increase in bleaching would also quite possibly increase the rate of accumulation of CO2 in the atmosphere, because coral reefs act as a large sink for CO2. By accreting calcium carbonate into their skeletons, they remove CO2 from the global cycle for very large periods of time. If large amounts of coral were to die as a result of temperature increases due to greenhouse warming, the rate of CO2 concentration increase in the atmosphere would climb.” (ENN) Of course, the same atmospheric result would be predicted if the corals died of simple starvation. The amount of carbon dioxide that living reefs effectively remove from the atmosphere is substantial: “It was found that in 5000 square meters of actively growing reef, between 2.4 and 9.7 tons of carbon dioxide was pulled from the water annually.” (quoted from Ocean Chemistry Division, NOAA) The active functioning of the living marine ecosystem is the main engine on this planet that is capable of removing CO2 from the atmosphere. It is called the “biological pump.” Destroying or “eliminating” large quantities of marine life therefore has the unavoidable consequence of atmospheric regression to higher CO2 levels. To make a long story short, atmospheric CO2 levels have had a few ups and downs over the history of the planet. Beginning with an essentially pure CO2 atmosphere, the evolution and accumulation of living organisms was associated with the lowering of CO2 levels. The main credit is generally given to the plants -- photosynthesis drawing CO2 from the air and incorporating it into the plant tissue and then the food chain. Today we’re very concerned with the rates of carbon-fixation achieved by plant life, and countries are hoping to earn “CO2 credits” by allowing forests to grow. Until very recently it was believed that terrestrial plant life removed more CO2 from the atmosphere on an annual basis than does marine plant life. Recently, however, based on satellite data, the conclusion has been reached that marine photosynthesis may outpace terrestrial photosynthesis in gross annual amount of carbon fixation. Regardless, marine photosynthesis naturally carries a much higher atmospheric significance than does the terrestrial variation. This is because, to quite a large extent, marine phytoplankton is consumed by SHELL-FORMING organisms (sometimes a succession of them)...which greatly amplifies the “atmospheric protection service” provided. (The shell formers effectively remove more CO2 from the water while constructing calcium carbonate; this is in addition to making use of the carbon originally fixed by the phytoplankton for somatic growth and energy requirements, as other fish do.) Viewed from this angle, sea plants become much more valuable than terrestrial plants as atmospheric “protectors.” It’s not only the nourishment of reef-building corals that provides “a large CO2 sink,” it is also the nourishment of every snail, mollusc and barnacle in the ocean, that works toward this end. Beyond those, there are billions of microscopic shell formers in the sea as well, all performing the same “atmospheric protection service.” Life evolved first in the sea, and the major part of the “CO2 correction” was accomplished by sea organisms before terrestrial life became well established. More than one strategy was used, photosynthesis was the first, but the tendency of phytoplankton consumers to form shells made of calcium carbonate was ultimately the more significant and effective CO2 removal method. Nothing comparable to this “shell building” evolved on land, therefore the sea has always taken the main responsibility for “adjusting” down the atmospheric CO2 level. Rather than incorporation of the CO2 into the food chain (where much or most of the time it is released again in fairly short order, in gaseous form through respiration or decomposition of organisms), this shell-building strategy effectively shunts carbon from the atmosphere into the mineral department. Uninvolved in the food web, carbon that follows the “sea shell” route ultimately remains locked up in stone for a very long time. Major portions of the earth’s crust are now composed of stone that originated in this manner - millions of years later the atmospheric carbon removed by the tiny sea creatures to build their shells is still “out of circulation.” Regarding the “carbon credits” that “developed” countries are wanting for forest growth, it is well acknowledged that only young, actively growing forests are really good carbon sinks. Old growth forests tend to reach an equilibrium between carbon fixation by new growth and CO2 release by death and decay. (And only a relatively very small fraction of terrestrial plant life ever becomes locked away in “fossil fuels.”) So, in terrestrial systems, old growth forests do not achieve a very impressive net CO2 removal rate. How does a comparable marine ecosystem look? How much CO2 removal is accomplished by an “old growth” marine ecosystem? The richer and more abundant the lifeforms in a marine system, the faster will be the accumulation of all sea shells, with the resulting expansion of coral reefs, formation of sandstone, limestone and sandy beaches. The shunting of carbon into the mineral department remains at a very high rate while the “old growth” marine ecosystem becomes stabilized by whatever other factors ultimately limit it. Like the terrestrial system, the living marine organisms might eventually saturate their ecosystem in the sense that CO2 release ends up equalling CO2 “fixed” into the food chain -- the significant difference in the aquatic system, however, is that the constant shunting of carbon into the mineral department is “permanent” and “ongoing” -- and that is the real key to why earth’s creatures rely on the ocean for the “sustainability” of life itself. Never before in the history of the planet did the land animals successfully pull the bulk of the sea animals from the ocean -- “fishing,” we like to think of it as a long established and “sustainable” tradition, but the fact is that fishing is a very recent development in geological time on this planet. (And our “nutrient” replacement strategy, basically amounting to coastal sewage contributions...has been an utter failure.) “Fishing” has had the ultimate effect of undercutting the very foundations of life in the sea, not only the coral reefs, and another unfortunate spin-off effect of this has been the slowing down of the “biological pump” in the ocean, the living planet’s only strategy for removing CO2 from the atmosphere. We might do a lot better to issue "carbon credits" for "feeding the fish." It is very interesting that the beginning of the current rising trend in CO2 predates the industrial revolution, but does not predate commercial fishing and whaling. It is also intriguing that a slight hesitation in the CO2 upswing coincided with World War II...which was not a time of less fossil fuel consumption (possibly it was increased?), but was a few years when fishing essentially came to a halt in the North Atlantic. For “whoever” was trying to remove CO2 from the planet’s atmosphere, however, the wartime presented a bit of a respite, and a wee gain was made briefly (CO2 levelled and dipped slightly during the war - see graph below - naturally the records from the last 100 years are the most accurate, and they show a steady upward course over the entire last century, with only the one little "glitch" near the middle. After 1945 the steady CO2 climb was resumed. This may look like a "small thing" but a really good explanation needs to try to account for these sorts of details. Actually, this writer was not the first one to notice the little glitch, scientists arguing against fossil fuel consumption as the cause of rising CO2 pointed it out first, and they are right, it's a little inconsistency that's hard to explain.) 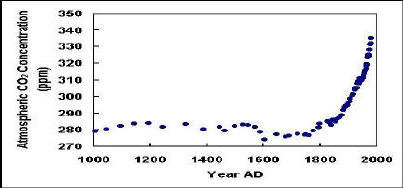 The creatures involved in driving the “biological pump,” the living sea creatures,
experienced more success at their job during the wartime, while the fishermen were
otherwise engaged. Fish stocks also made remarkable gains at that time. Given a chance,
the very resilient life in the ocean could still get to work and correct the rising CO2....but
they cannot do it at this point without our co-operation. We need to drastically reduce
fishing and make some deliberate, sensible efforts to nourish the life that still remains in the
sea. The creatures involved in driving the “biological pump,” the living sea creatures,
experienced more success at their job during the wartime, while the fishermen were
otherwise engaged. Fish stocks also made remarkable gains at that time. Given a chance,
the very resilient life in the ocean could still get to work and correct the rising CO2....but
they cannot do it at this point without our co-operation. We need to drastically reduce
fishing and make some deliberate, sensible efforts to nourish the life that still remains in the
sea.
Many scientists maintain that the only real way to save the corals is to reverse global warming.......a closer look reveals the insight that the only way to solve EITHER problem may be to REVERSE FISHING! -- Theodore Roosevelt Debbie MacKenzie REFERENCES Abney, Michael A. & Joe E. Clemons. Phenotypic plasticity in the reproductive life-history traits of Caribbean gambusia from the Cayman Islands, BWI ( from: http://www.animalbehavior.org/ABS/Progran/Past/Morehouse_00/absAL.html ) Barnes, R. S. K., and R. N. Hughes, An Introduction to Marine Ecology (third edition). Cambridge: University Press, 1999 Brown, B. E. Coral bleaching: causes and consequences. Coral Reefs 16:S129-S138, 1997 Chadwick, Alex. 2001. The Treasured Islands of Palmyra. National Geographic Magazine, March 2001. Fitt, W. K. et.al. 1993. Recovery of the coral Monastrea annularis in the Florida Keys after the 1987 Caribbean “bleaching event.” CORAL-REEFS 1993, vol. 12, no. 2, pp 57-64 Glynn, P. W. 1996. Coral reef bleaching: facts, hypotheses and implications. Global Change Biology 2: 495-50 Hart, A. M., Russ, G. R. Response of herbivorous fishes to crown-of-thorns starfish Acanthaster planci outbreaks. Age, growth, mortality and maturity indices of Acanthurus nigrofuscus. MEPS 136:25-35 (1996) Hayes, R. L. and Goreau, N.I. 1998. The significance of emerging diseases in the tropical coral reef ecosystem Revista de Biologia Tropical 46 Supl. 5: 173-185 Hoegh-Guldberg, O. 1994. The population dynamics of symbiotic zooxanthellae in the coral Pocillopora damicornis exposed to elevated ammonia. J. Pacific Science 48:263-272 Hoegh-Guldberg, O. 1999 (or 2000?, online doc not dated). Climate Change, Coral Bleaching and the Future of the World’s Coral Reefs. - accessed through Greenpeace’s Australian website: www.greenpeace.org.au ISBN 90-73361-52-4 Hudson, J. H., Hanson, K. J., Halley, R. B., Kindinger, J. K., 1994. Environmental implications of growth rate changes in Montastrea annularis: Biscayne National Park. Florida Bull. Mar. Sci. 54(3): 647-669 Humm, Harold J. and Susanne R. Wicks. Introduction and Guide to the Marine Bluegreen Algae. New York: John Wiley and Sons, 1980 Idso, Craig D. and Keith E. Idso. “Coral Reefs: Doomed by Carbon Dioxide?” from www.co2science.org 1999 Idso, Craig, D. and Keith E. Idso. “Disease as a Cause of Coral Bleaching - Summary” from www.co2science.org 2001 Lassus, Patrick et. al. Harmful Marine Algal Blooms - Proceedings of the Sixth International Conference on Toxic Marine Phytoplankton, October 1993, Nantes, France. Andover, UK: Intercept Limited, 1995 Lough, J. M., and Barnes, D. J., 1997. Several centuries of variation in skeletal extension, density and calcification in massive Porites colonies from the Great Barrier Reef: a proxy for seawater temperature and a background of variability against which to identify unnatural change. Journal of Experimental and Marine Biology and Ecology 211: 29-67 McCook, L. J. 1999. Macroalgae, nutrients and phase shifts on coral reefs: scientific issues and management consequences for the Great Barrier Reef. Coral Reefs 18: 357-367 Merrett, Nigel R. and Richard L. Haedrich. Deep-Sea Demersal Fish and Fisheries. London: Chapman and Hall, 1997 NOAA. 1998 (on-line). “The Extent and Condition of U.S. Coral Reefs” by Steven L. Miller and Michael P. Crosby. NOAA’s State of the Coast Report. Silver Spring, MD: NOAA URL: http://state-of-coast.noaa.gov/bulletins/html/crf-08/crf.html Ogden, John C. and Elizabeth H. Gladfelter (eds). Coral Reefs, Seagrass beds and Mangroves: Their interaction in the coastal zones of the Caribbean. UNESCO Report on a workshop, 1983. Pecheux, Martin. “Review of Coral Bleaching” (on-line) 1992. accessed at http://www.essi.fr/~sander/articles/Misc/Coral-Reef.html Polunin, Nicholas V. C. and Callum M. Roberts (eds). Reef Fisheries. London: Chapman & Hall, 1996 Sebens, K. P. 1994. Biodiversity of coral reefs: What are we losing and why? Am. Zool. 34(1): 115-133 Sherman, Kenneth, Lewis M. Alexander, and Barry D. Gold (eds). Food Chains, Yields, Models and Management of Large Marine Ecosystems. Oxford: Westview Press, 1991 Wilkinson, Clive -- online status of reefs report, link from NOAA Woodard, Colin S. Ocean’s End. New York: Basic Books, 2000 |