- How does “nutrient” cycling work on the
reefs? - How do these systems respond to changes in “nutrient” levels? - What is the significance of the form (solid/liquid) of the nutrients? - What is the ecosystem response to the removal of organic biomass by
fishing? -Are fish equivalent to “nutrients?’ - Might their removal be equivalent to
“nutrient” loss? - Can fishing negatively affect “primary production?" - How do the
changing trends in coral reef ecosystem compare with those in other marine systems?
1. Coral Reef Ecosystem dynamics.
At first glance, coral reef ecosystems seem to present something of an incongruity.
A healthy coral reef is a diverse, highly “productive” community of marine organisms,
thriving in exceptionally “nutrient poor” waters. “Productive” refers to the relatively high
amount of “carbon fixation” that takes place in these systems...measurements and
calculations have been made to show that by this measure, coral systems rank among the
most productive of marine ecosystems anywhere. And this is accomplished in the relative
absence of dissolved nutrients (N and P) in the clear, “oligotrophic” water. Rather than
being a “poor” system, however, the coral reef is a “rich” living system that manages to
have (and keep) it’s nutrients largely tied up in solid, living matter. The standing stock of
reef fish represents a significant nutrient resevoir for a these systems.
The secret to the success of the coral reefs is commonly believed to be the “tight
recycling of nutrients” in the system, particularly in the corals, in which tiny plants and
animals live together in a symbiosis that conserves key nutrients quite effectively. Algae
(zooxanthellae), living inside the tissues of the cnidarian host, harness energy from sunlight
and “fix carbon” by photosynthesis. Energy from this source is provided to the polyp host
in return for exclusive access to the waste-nutrients produced by the host. These wastes (N
and P) function to fertilize the algae. The most significant, and apparently “limiting”
nutrient in the picture is fixed nitrogen, a critical element in the construction of all proteins.
The symbiotic arrangement allows the partners to avoid the loss of fixed N to the water, a
process that normally occurs in both free floating algae and solo-living marine animal
lifeforms. In larger marine systems, overall fixed N is essentially passed back and forth
between the plant and animal compartments of the community - corals have evolved a way
to capture and complete this loop inside their own special symbiosis. It’s a micro-model of
what happens in the bigger picture, and the same principle applies in the more “nutrient”
rich systems of the temperate and polar zones. Plants and animals perpetually passing the
precious ball (fixed N) back and forth - it’s a common theme in all living systems on the
planet.
It is definitely an efficient plan for conserving nutrients, but the coral-algae
symbiosis cannot live on sunshine, CO2 and water alone. A net input of N and P is still
essential to maintaining life and growth, and all corals have feeding strategies in addition to
deriving energy from their plant partners. The partners have the ability to extract dissolved
bio-available N from the water at low ambient levels, and also they capture and consume
microscopic prey (zooplankton) as well as bacteria and particles of edible detritus that
come into contact with their mucus layer. In short, corals also need to eat to live.
Like many organisms, corals are adept at storing food-energy for the lean times.
Under favorable feeding conditions they can become quite “fat.” This is important, since
the food sources listed above may not be consistently available. Of the dissolved forms of
fixed N, ammonia is by far the most easily available for uptake from the water by the
symbionts, and this apparently is true for phytoplankton in general. Since living fish
constantly excrete ammonia from their gills....it suggests that removing major amounts of
fish from the system “might” ultimately deprive corals of needed nutrients. That’s the
hypothesis.
So, the coral reef ecosystem as a whole is characterized by a relatively low net
exchange of nutrients with the surrounding areas. How much do we know about the overall
input-output patterns, or “nutrient balance” in the system? The capture of plankton from
oceanic water passing over the reefs is thought to be one important input route, although
this water is characteristically very “low” in nutrients. Another significant input is the
contribution of fixed N by the blue green algae - this group of organisms does well in
tropical water since they can out-compete other N-limited algae forms, having the
advantage of “making their own fertilizer.” As described in a basic marine ecology text:
“Atmospheric nitrogen is also fixed by blue-green algae such as Calothrix crustacea,
which occurs on intertidal reef flats in the Pacific as a thin, mono-specific film (it also
occurs in other reef habitats in different growth forms). Calothrix can fix nitrogen at the
rate of 1.8 kg/ha/day -- two to five times the rate achieved by fields of lucerne or alfalfa.
Fixed nitrogen enters the food web through at least three routes: (1) the blue-green algae
are consumed by herbivores, especially by certain fish with low assimilation efficiencies, so
that the water over the reef gains nitrogen via the fish faeces; (2) in areas subject to strong
wave action, large peices of the Calothrix film are dislodged and washed over the reef,
where they will be available to consumers; (3) Calothrix releases about 50% of is fixed
nitrogen into solution, from which it may be taken up by other autotrophs.” (Barnes and
Hughes, 1999, page 140).
Besides the capture of oceanic plankton, and the use of N fixed by the blue-greens,
many coral reefs derive significant nutrient input from terrestrial sources. (Although a
normal feature in many undisturbed areas, the “mangrove-seagrass-coral reef” type of
system does not support all coral communities. Undisturbed, the atoll reefs appear able to
sustain themselves very nicely without a current direct route for terrestrial source nutrients.
Efficient recycling is the key.) Elsewhere, in the “mangrove-seagrass-coral reef” systems,
the mangroves grow at the shoreline, tolerant of relatively high dissolved nutrient levels,
very “productive,” and providing habitat for a wide range of other organisms. The
mangrove environment makes use of large amounts of dissolved nutrients, also significant
denitrification takes place there, so the natural effect of the mangrove growth is a
significant drop in nutrient levels in the water. Seaward of the mangroves are the seagrass
beds, another highly productive area that also removes a significant amount of dissolved
nutrients from the water. Seaward of the seagrass beds are the coral reefs, described as
“oligotrophic systems least tolerant of nutrient enrichment.” Mangrove forests and seagrass
beds both provide ideal feeding and shelter for a variety of juvenile reef fish, and they are
well known to function as “nursery areas” for these. From “Coral Reefs, Seagrass Beds
and Mangroves” (UNESCO, 1983):
“The sheltered nature of these areas also contribute to make them important as
nurseries. Mangrove areas thus export protein to coastal areas in the form of aquatic
organisms that use the mangrove areas for their early development and then migrate
offshore. Well known are the massive migrations of mullets and shrimp from these areas.
This is high quality protein that links mangroves directly to other coastal systems like coral
reefs, seagrass beds, and ultimately to man.” (Bak in Ogden and Gladfelter, UNESCO
Report, 1983).
The function of the mangrove and seagrass areas could be described as converting liquid
nutrients to solid nutrients, prior to their export to the coral reef....in other words, organic
“runoff” is converted to fish flesh by this system. And the “fish flesh” type of “nutrient
enrichment” seems to be quite acceptable to the reef ecosystem...so “oligotrophic systems
least tolerant of nutrient enrichment” isn’t precise enough, it’s only liquid forms of nutrient
enrichment that are “intolerable.” In contrast, the solid, swimming fish form of “nutrients”
has traditionally been very well “tolerated” by the reefs.
One other natural route of net nutrient input into these tropical ecosytems may once have been significant.
Large migratory animals, like the great whales, that travel long distances to the temperate
zones to feed; these animals spend part of the year in the tropics. Not feeding, but
subsisting on stored energy from their northern excursions, the whales are in the warm
water to give birth and nurse their young. Therefore, on occasion, whales must die naturally in the tropics. If their
corpses come to rest within scavenging distance of the coral reef fish, that event
must be considered a significant nutrient input, and it is of a type that is natural and
acceptable, non-damaging to the ecosystem. And a key feature of the “acceptability” of this
“nutrient enrichment” is its solid form. (If the dead whale came to rest beside a coral reef
and was ultimately consumed bit by bit, by the organisms living there, the sea water would
remain clear, and life in the community would basically go on as before, just a whole lot of
little consumers growing a bit fatter....on the other hand, if the dead whale had first been
put through a “blender” and then the nutrients were poured over the reef, a rapid bloom of
free-floating phytoplankton and an increase in sedimentation and a decrease in sunlight
hitting the corals....an unhealthy mess, a quite different result would be obtained.)
One theme that emerges from the story is this one:
It’s the FORM of the nutrient input that is highly significant to aquatic systems. It
actually comes down to the important advantages of “SOLID” vs “LIQUID” food.
We are often reminded that coral reefs are “intolerant of nutrient enrichment” and normally
“nutrient poor.” More accurately it should be stated that they are “intolerant of LIQUID
nutrient enrichment” and normally “LIQUID (dissolved and particulate) nutrient poor.” As
the previous discussion has (possibly) made clear, most of the “normal” or naturally
acceptable forms of nutrient input to reef systems are SOLIDS. The most acceptable (and
significant) form of terrestrial-source nutrient input to the coral reef systems arrives in the
form of living, swimming fish that have ingested nutrients nearshore, grown larger and then
migrated off to the reef. The fixed N contributed by the blue-green algae enters into the
web to a large extent via fish feces, or by organisms eating the solid algae mats directly.
The oceanic plankton captured basically also represents tiny solid forms, dissolved nutrients
playing only a very minor role in the larger “input” picture of the reef.
What about natural “losses” of nutrients from coral reef systems? Some nutrients are
inevitably swept away in the seawater, and some will undergo denitrification in the seabed.
Accurate quantification of the amount of fixed N lost by the system in these ways is really
not possible with the state of today’s knowledge. Therefore, the annual amount of “extra”
fixed N/protein in the reef system -- the amount that could safely be removed without
diminishing the system overall -- is unknown, but seems unlikely to be a very great amount.
The other obvious mechanism of “nutrient loss” from tropical ecosystems is fishing. The
removal of fish from these systems by human fishing most likely represents the biggest net
loss of nutrients. Large quantities of “solid nutrients.” The argument offered in the
temperate zones that “the food web can always replace the fish that have been removed
because the ocean contains a vast pool of bioavailable nitrogen”....well, that particular line
of reasoning will obviously not go far in the tropics, since it is essentially referring to some
great reserve of dissolved (liquid, non-living) nutrients. It is clear that the tropical fish
WERE a major fraction of the “nutrient pool” themselves. Is it likely or possible that the N
fixing activity of the blue-green algae can keep pace with human fishing removals? If so,
they would need to “pick up the pace” very significantly, and fix N at a greater rate than
they did during the pre-fishing millenia of the reefs’ existence. Have they done this? It
seems not, considering the current depleted state of life on many reefs....many will say
that’s only because they have been “overfished”...but that’s only a term referring to recent
fishing intensity, a more credible argument may be that all-fishing, over the recent centuries
of human exploitation, has greatly reduced the overall quantity of life in the reef systems.
It’s because we are the “unnatural predator,” the one who takes but does not give back
(anything useful)...and that is part of the essence of the whole problem with disappearing
marine life today. Mass coral bleaching (death of clean water corals by food starvation
during warm spells) is a truly new and ominous phenomenon, and it’s most likely to be merely the ultimate
end result of all the fishing that humans have done in the tropics.
Regarding the importance of the form (solid vs. liquid) in which nutrients exist in the coral
reef system, and indeed in all marine ecosystems, I would like to elaborate on this point a
bit further. It seems that fish assimilate on average about 10% of the energy and nutrients
in the food that they consume, and the remaining 90% they excrete rather promptly back
into the environment. In discussions of this point with marine biologists, one
gets the impression that the 90% that’s not assimilated is shunted back to the
bottom of the food web for recycling, in effect it’s transformed into the equivalent of
human sewage. But this seems to be somewhat less than accurate, because when our
sewage hits the ocean it’s essentially liquid.
Admiring the fish in an aquarium store recently, I became very interested in the
question: “why do fish excrete solid feces?” Not a terribly extensive survey to be sure, but
all of the fish that I observed, both the freshwater and salt water species, were defecating
what can only be described as “formed stool,” which was sinking to the bottom of the
tanks. Now why would this be the case? Land animals resorb the water from their feces
before excretion presumably because we are conserving water. But for the purposes of
digestion in all animals, the way to efficiently absorb nutrients from one’s food is to
convert it to liquid form in the gut. So...if fish do not have a great need to conserve water,
why don’t they just excrete their feces in liquid form? After all, that would be the quickest
way to send the nutrients back to the “bottom of the web” to be recycled. If they did, the
nutrients would be better dispersed throughout the water column and more likely to be
quickly taken up in the process of photosynthesis. But instead, fish excrete a solid stool that
sinks. By keeping the waste product in a solid form, the nutrients remain more available for
consumption by a wider range of scavengers, and relatively less available to the
phytoplankton. Even the tiniest sea animals, the zooplankton, apparently excrete what is
termed “fecal pellets” (which sink). One underlying principle in aquatic nutrient recycling
seems to be: “minimize the liquid input, as much as possible keep edibles in a solid form.”
This may be a reflection of the low tolerance for liquid nutrient input in living aquatic
ecosystems. It appears that fish excrete ammonia and urea in liquid form, both are forms of
fixed N that are very easily available to the phytoplankton, and are therefore perhaps
“intended” for their use...and the remainder of the fishes’ food waste is packaged in a solid
sinking form, intended for recycling at a “lower level” but not necessarily the very
“bottom” of the web. This strategy may be an important part of the overall design to
preserve the balance between the many organisms living in the system.
The consumption of another’s feces, (a concept most repulsive to us), is a routine
part of nutrient cycling in nature. Even carnivores (once we give them that label, do we
only imagine them consuming flesh?) are known to consume feces. One example is the
codfish in the North Atlantic - they are known to consume seal feces. A parasitic worm
takes advantage of this fact and plans its life cycle accordingly -- from our point of view it’s
“distasteful,” we hate to see the worms in the cod fillets -- but from the point of view of
nature, it’s just the seals putting leftover “nutrients” into the proper recycling bin. And it is
just one example of a natural recycling route whose success depends on the fact that the
nutrients are maintained in solid form. Many similar feedback loops are likely completed
between other groups of marine organisms without the basic nutrients returning each time
to the “bottom” (phytoplankton) to be recycled.
Of all the variations on the marine ecosystem theme, coral reefs appear to be the
example that best illustrates the intolerance to excessive amounts of liquid nutrients. This
intolerance is a characteristic of all aquatic systems, but the fact that this feature is taken to
such an extreme in the case of the coral reefs, helps to educate us about the underlying
principle, one that is valid in the oceanic system overall -- The form, solid or liquid, in
which nutrients are added to marine ecosystems is of critical importance in
determining the likelihood that the system will be able to assimilate them and
incorporate them into the “web of life.” Too much liquid causes an imbalance and
sickens the system, and mechanisms will be used by nature in those cases to get rid of the
excess, unusable nutrients. The capacity of these systems to make use of solid food input
far exceeds their capacity to assimilate liquid food. This is one key reason why our current
pattern of removing solid fish from the sea and “replacing the nutrients” with
organics-loaded liquid runoff...just does not work. Too quickly, this approach saturates the
ability of the system to accept liquid food, and triggers the “get rid of it” strategies. “IF” we
changed our behavior and started to “feed the ocean” with solid food, we could expect to
see a far healthier result....
Another theme in this discussion is an attempt to dispel the myth that the ocean is
currently “nutrient overloaded.” Reassurances given that “total fixed N in” (largely runoff)
exceeds “total fixed N out” (fishing and natural routes) don’t hold up, once the comparative
value of solid vs. liquid nutrient forms is factored in. (And beyond the form, the location of
the N-input (coastal only) is inappropriate, if it really is “intended” to make up for the
fishing-induced losses (significant amounts from offshore)).
....but this is leading away from the main topic in this essay...so, back to the coral reefs. What
is happening? How much to we know?
2. EXACTLY HOW DO CORALS RESPOND TO CHANGES IN NUTRIENT
LEVELS?
“Too much” liquid nutrient input clearly seems to damage them, the
“eutrophication” pathology picture has been fairly well examined and described in the
literature. Has the possiblity of the other extreme (“oligotrophication?”...is that a word? --
excessive nutrient depletion, anyhow)...has that one been investigated? (Even if it were,
absolute measurements of ammonia and nitrate levels are unlikely to be enough to tell the
tale -- they might remain unchanged and “nutrient depletion” of the system overall could
still be a reality due to the fact that so many “fish”(aka “nutrients”) are missing. At the very
least it would probably be necessary to average measurements over the year -- the lows
may not be getting any lower, it might just be the high points that are coming down.)
First, “excessive nutrient loading,” eutrophication caused in reef systems near to human
population centers. Unnaturally large amounts of liquid nutrients (dissolved and particulate)
run off into coastal waters, from sewage, fertilizers, aerial deposition and sediment/soil
erosion. These consequences of human activity clearly threaten corals. Eutrophication is
commonly defined along the lines of “an increase in the rate of supply of organic matter to
an ecosystem.” One of the obvious signals that an aquatic system is eutrophied is an
anomalously high increase in phytoplankton biomass. The growth of macroalgae also
seems to be favored under these conditions. How exactly does this hurt corals? Water
becomes increasingly turbid as a result, so corals receive less light, also the rate of
sedimentation increases as phytoplankton die and sink, decomposition can result in oxygen
depletion....in short, an unhealthy chain of events is set in motion which threatens the
continued health and dominance of the corals. Overgrowth of algae, and possibly also a
disproportionate increase in benthic filter feeders, which can also displace corals and
accelerate bioerosion of the reef - these also can be part of the picture of a reef degraded
by eutrophication....or at least that has been the commonly accepted thinking. Recent
research, however, on the shift from coral to macroalgal dominance (a common picture in
reefs experiencing eutrophication) indicates that this effect, presumed to be a direct impact
from liquid nutrient enhancement, may not be exactly as imagined. Apparently, accomplishing the "shift" also requires that the population of herbivorous fish be
significantly depleted. (McCook, 1999)...so "fishing" does factor into the "shift to algal dominance," it's not effectively driven by "nutrient enhancement" alone.
But at the same time, nutrient enhancement alone does have the potential to harm corals.
Olivieri (1997) describes some of the fine points of “eutrophication,” including how excess
nutrients have a harmful effect on coral by overstimulating the growth of zooxanthellae to a
point where their “uncontrolled” growth disrupts the normal function of the coral:
“Eutrophication also operates at the zooxanthellae level. Excess nutrients increase
zooxanthellae growth, which, counterintuitively, is not beneficial for the coral host. Under
natural conditions zooxanthellae populations are fairly constant and nutrient-limited,
particularly by nitrates, within the coral host. With excess nutrients the zooxanthellae
population grows uncontrolled and the balance of the nitrogen-carbon fluxes between the
coral host and zooxanthellae is disrupted, resulting in a reduction of calcification and
weakening of the coral calcareous skeleton.” (Olivieri, 1997)
But exactly how much is too much?
Experimental studies of increasing ammonia levels available to corals have shown that
initially an increase results in accelerated normal growth of zooxanthellae and also an
increase in growth of the polyp host. Above a certain threshold, however, the presence of
more ammonia in the water stimulates a disorganized growth pattern in the zooks - and this
is likely the point where the enrichment becomes disadvantageous for the coral as a whole.
But a general statement that nutrient enrichment interferes with the growth of corals
obviously cannot be made. They are certainly not completely “allergic” to “nutrient
enrichment.”
Ove Hoegh-Guldberg in 1994 reported a study entitled “The population dynamics of
symbiotic zooxanthellae in the coral Pocillopora damicornis exposed to elevated ammonia”
in J. Pacific Science 48: 263-272. :
“The presence of low concentrations of NH4Cl (0.2 µM) did not affect (compared to
corals growing in NH4+-stripped seawater) either division synchrony or growth rate.
Exposure to higher concentrations of NH4Cl (20 mM or 50 µM), however, affected the
population dynamics of the zooxanthellae residing in P. damicornis. Zooxanthellae in corals
exposed to 20 µM NH4Cl had mitotic indices (the percentage of the total cells dividing)
that were two to three times higher than the mitotic indices of zooxanthellae in control (0.2
µM) corals. Although the division of zooxanthellae was still phased in corals exposed to 20
µM NH4Cl, there were many more cells dividing out of phase as compared to control
corals. The division of zooxanthellae in corals exposed to 50 µM was not phased. The
calculated growth rates of zooxanthellae exposed to 20 µM or 50 µM NH4Cl were higher
than those representative of zooxanthellae living in control corals, although the growth rate
of both carbon and nitrogen pools were lower in 50 µM as compared to 20 µM NH4Cl.
These data support the conclusion that the population dynamics of symbiotic zooxanthellae
within P. damicornis are affected by concentrations of NH4Cl in seawater that are equal to
or higher than 20 µM, and that 50 µM NH4Cl concentrations may be toxic to some extent.
These data taken in isolation, however, do not constitute an effective test of the hypothesis
that zooxanthellae are limited by the supply of NH4Cl under ambient conditions, and
further emphasize the importance of enrichment studies concentrating on growth and
nitrogen incorporation rates measured for the entire symbiotic association.”
Maybe not. But it is an interesting hypothesis, “that zooxanthellae are limited by the supply
of NH4Cl under ambient conditions”....since important “suppliers” are fish.
What would a normal ammonia level be on a coral reef?
“Nitrogen sources for a coral include both organic nitrogen from ingested food and
inorganic nitrogen in the form of nitrate and ammonia...Ambient levels of these nutrients
are usually low: 1.0 uM nitrate and .02-0.8 uM ammonia at most reef sites except for inner
bays where measurements are twice these values...(corals) are functioning as oligotrophic
phytoplankton. In addition, at ambient concentrations of these two sources of inorganic
nitrogen, the rate of uptake of ammonia is twice that of nitrate. This suggests that ammonia
is probably the more significant of the nitrogen sources as is true of many marine
phytoplankters.” (Gladfelter, 1983)
In the same report she included the observation that: “Lewis and Smith (1971)
demonstrated that the amount of C14 incorporation by animal tissue as alanine was
increased if the medium was spiked with 5 uM ammonia.”
So, the ambient concentration of ammonia in reef water is generally under 2 uM, spiking it
to 5 uM has been shown to result in increased growth in the polyp, and even increasing it
10 times to 20 uM seems to result only in an increased rate of normal coral growth. Only
ammonia levels higher than 20 uM seem to result in increasingly disorganized and
deleterious growth patterns. This fact has been noted by other sources as well, for example,
from http://www.marinebiology.org/coralbleaching.htm , “Rather than causing coral
bleaching, an increase in ambient elemental nutrient concentrations (e.g. ammonia
and nitrate) actually increases zooxanthellae densities 2-3 times. Although
eutrophication is not directly involved in zooxanthellae loss, it could cause secondary
adverse effects such as lowering of coral resistance and greater susceptibility to diseases.”
........ The effect of INCREASING the density of zooxanthellae (thereby deepening the
color of the coral!) is directly opposite to the scenario involved in recent mass coral
bleaching events, the density of the algae is well known to DECREASE prior to their loss
by the host. This rather strongly suggests that an insufficient supply of their preferred N
source (“ammonia”....stuff constantly excreted through the gills of all living fish...) is a
more likely trigger for the bleaching problem. It looks like the coral’s tolerance for
ammonia is significantly higher than today’s “ambient levels” on reefs -- not surprising
considering their long history of coexisting with far higher “ambient levels” of LIVE FISH
in the sea.
One other aspect of the eutrophication problem is the (good fortune?) that it really is only a
nearshore problem. The addition of large quantities of liquid nutrients to aquatic systems
triggers the compensatory mechanisms that were built into the system to counteract them.
(Phyto bloom, with sedimentation, death and decomposition, denitrification intensifying as
nitrate levels increase. This scenario appears to be a built in homeostatic mechanism in
aquatic systems. And it works, the excess nutrients will be removed before they move too
far from shore.)
But corals are dying in areas with “pristine water,” not affected by eutrophication...that’s
why the offshore deaths seem so mysterious, and their coincidence with the annual high
temperature has lead to the suspicion by many that “water temperature” or “climate
change” is the primary cause. Indeed, media reports of the problem conveyed to the
general public relate the coincidence of coral bleaching and warmer water temperatures as
if it were a simple case of cause and effect. Scientists studying the problem know better
than this, but the general public hasn’t got a clue....
3. “OVERFISHING” IS A MAJOR THREAT TO CORAL REEFS
This observation is
commonly made today. But exactly what is the nature and consequences of this threat?
How does fishing have a negative impact on coral reef communities?
1. Local community short-term impact of removing a particular species. Usually described
as a “shift” in species composition rather than being seen as a “loss.”
“Fishing activities are often aimed at a particular trophic level within a community or at one
particular species. By removing either predators or competitors from a system fishing can
confer a competitive advantage on species that were previously constrained.” (Barnes, p
242)
“Overexploitation affects the vast majority of the world’s reefs...At a minimum, overfishing
results in shifts in fish size, abundance, and species composition within reef communities.
Evidence suggests that removal of key herbivore and predator species may ultimately affect
large-scale ecosystem changes. For example, removal of triggerfish has been linked with
explosions in burrowing urchin populations, their prey, who subsequently accelerate reef
erosion through feeding activities.” (World Resources Institute, online article “Threats to
coral reefs, coral ecosystems”).
Unusually large outbreaks of coral-eating “Crown of thorns starfish” have damaged reefs in
quite a few places in recent years, including the Great Barrier Reef of Australia. This seems
to be a suspected result of fishing removal of their predators -- “overfishing” the
carnivorous reef fishes. And that much is not hard to believe.
The removal of herbivores is similarly seen as resulting in increased algae presence on
reefs: “In the Caribbean, decades of overfishing has led, in many places, to very low levels
of grazing fish species. Because of this, herbivorous sea urchins (a nonburrowing species)
have played an increasingly important role in keeping down algal growth. In the early
1980s, huge numbers of these urchins succumbed to disease. Without grazing fish or
urchin populations, and spurred on in many areas by organic pollution, algae quickly
dominated the reefs, inhibiting coral settlement and sometimes overgrowing living corals.”
(WRI)
(The removal of herbivores plus the addition of organic pollution are seen as both
contributing to the increased growth of algae on the reefs. Entirely possible, of course.)
2. Destruction caused by fishing methods
Physical damage to habitat and non-targetted
species. This is a major concern in many heavily exploited reef systems. “Blast fishing,
fishing with cyanide and other poisonous chemicals, muro-ami netting (pounding reefs with
weighted bags to scare fish out of crevices), and in deeper waters, trawling directly damage
corlas. Because these methods are generally nonselective, large numbers of other species,
along with undersized target species, may be swept up in nets or killed by poisons or
explosives in the process...As not all fishing methods are destructive, this is less of a
widespread threat than overexploitation.” (WRI)
3. Fewer fish.
“Overfishing” has one obvious and direct impact on reef fisheries: less fish
become available for human harvesting and consumption.
4. Reducing the standing stock of reef fish lessens the aesthetic appeal of the reefs thereby
possibly damaging their value to the tourism industry.
5. Occasionally, one will encounter the phrase “corals depend on fish” but it inevitably
seems to end up meaning that corals depend on fish to consume their algae competitors
and thereby maintain a more coral-friendly environment.
The prededing five points summarize what seems to be the usual thinking about how
fishing degrades or damages coral reef ecosystems. But there is one other
mechanism of harm wrought by fishing here, one not noticed because of its insidious
nature perhaps; it seems most likely that fishing removal of organic “biomass” leaves
behind a generalized nutrient deficit...in a system with a known practice of “tight
nutrient recycling.” Have we somehow forgotten to state the obvious:
“FISHING
REMOVES FOOD FROM THE SEA”...LOGICALLY RESULTING IN LESS
FOOD BEING AVAILABLE TO SUSTAIN THE LIVING MARINE SYSTEM?
Is there theoretical or proven evidence to support or to deny this particular
hypothesis?
“One of the universal processes inherent in all ecosystems is the recycling of matter.
Without this process life on Earth cannot be sustained, as most of the 30-40 elements
necessary for the growth and development of living organisms are in finite supply.”
(Barnes & Hughes, p 233)
Safe to say, similarly,.... “without this process life in the sea cannot be sustained?”
And regarding finite supply, fixed nitrogen is recognized as most often being the limiting
nutrient in marine systems. Therefore efficient and effective recycling of fixed nitrogen is a
basic essential requirement for the continuation of life, and scientific evidence for a real
balance between fixed N removed by fishing, and fixed N put back by humans...does not
exist. (In contrast, it is a simple thing to delineate the exact routes by which all natural
predators in the sea efficiently recycle the fixed N and other nutrients that they have
consumed during their lives. They seem to operate on the simple principle “keep most
nutrient ‘inputs’ in solid form” as this keeps them “in the game” and doesn’t trigger the
systemic decompensation that kicks in when too much of the total is reduced to liquid form
at one time. For best results, fertilize the phytos only lightly...)
Regarding coral reefs this is a typical explanatory statement:
“High production with very low available nutrients is explained by high levels of
nitrogen fixation plus very intense nutrient recycling.”
The understood meanings of the words, “nutrients” and “production,” might be
contributing to the confusion. Reference has already been made to the problem with using the
word “nutrient” to denote those existing in liquid form only, and how this obscures the
reality of how much “solid nutrient” has disappeared. The word “production” can also
cause confusion - it seems that it has a couple of noticeably different meanings.
“Primary production” in marine biology refers to the amount of carbon fixed by the
phytoplankton, basically the rate of conversion of CO2 to food. Expressed this way,
estimates of “productivity” on coral reefs might be something like “7,000
gC/m(squared)/year.” A lot more carbon fixation takes place on the reefs as compared to
the surrounding areas. The potential problem is when the assumption seems to be made
that such a “highly productive” system can afford to give up a lot of fish repeatedly....there
seems to be an implication that there should be no trouble in replacing them...just
“producing” more. A lot of carbon may be fixed, but a lot of it may be reconverted to CO2
via respiration of organisms...and a lot more may become rather permanently sequestered
in the seabed. It is not at all clear that there is any relationship between “grams of carbon”
fixed on an annual basis, and the potential for “sustainable” fisheries yield.
“Carbon fixed by photosynthesis on the reef is lost partly by offshore transportation but
most is accounted for by the metabolic activity of intermediate consumers. The
production to respiration ratio (P:R) therefore is close to unity and there is insufficient
fixed carbon left to support high, sustainable yields of large carnivores at the end of the
food chain.” (Barnes and Hughes p 141)
“Close to unity?”....meaning that most of it is used up in that manner (respiration). So only
a relatively small amount would be left over to invest in building new fish flesh?
“...the end of the food chain.” -- That is another unfortunate phrase, since it’s a reflection
of human linear thinking...of the sort that has gotten us into this kind of trouble in the first
place... “The food chain” concept may serve to distract us from really seeing “the food
cycle” -- a more accurate image being of a CIRCLE, with no “end” -- that’s really what the
recycling design was based on. The “food chain” is circular, not linear!
...Acknowledging this is hard for us to do however, since it is rather difficult to explain
how it is that humans participate in the marine “circle” of perfectly balanced give and take
-- it’s undeniable that that was the ecosystem plan that worked to sustain marine
communities for eons. (We like to think of ourselves as just another top predator, happily
glossing over the differences between ourselves, seals and sharks...) The balance of species
in the sea doubtless shifted many times, and even the “total biomass” was subjected to
natural fluctuations as the planet changed, but over time, and largely due to the N-fixing
work of the blue-green algae, a great wealth of organic “nutrients” was accumulated in the
sea. The most-valuable N was carefully conserved and recycled to the greatest extent
possible by the living organisms. And, any way you look at it, “Mother Ocean” is just not
looking particularly “wealthy” anymore...
“There are plenty of high-profile examples of marine species, such as whales, dolphins,
seals, turtles and seabirds, that are considered to be endangered. However, these
organisms tend to be at the top of the food chain, and so contribute little to the
productivity of the ecosystem. Of more concern are the abilities of human activities to
affect animals from lower trophic levels, the populations of which are considered, often
over-optimistically, to be more resilient to exploitation.” (Barnes and Hughes, p 252,
1999)
There it is again, “the top of the food chain”...as if there is nowhere to go from there,
except possibly “up” another notch to be consumed by humans (?). And what exactly is the
meaning to be taken from the word “productivity” in that sentence? There’s the myth
again, that the creatures at the higher trophic levels are not particularly important to the
functioning of the ecosystem as a whole. But the truth is that they were simply living there
and practicing “efficient recycling of nutrients,” just like all their neighbours at the “lower trophic levels.”
That quote was taken from a university level textbook on marine ecology published in
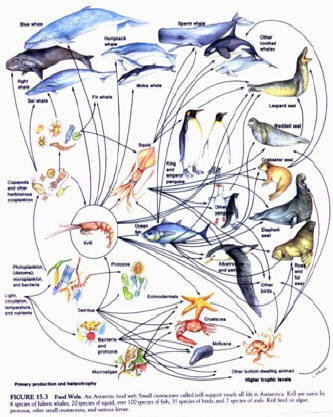
1999. Granted, a lot of current literature uses the term “food web” instead, and many
diagrams exist of “food webs” which are essentially a complex intertwining of many
upward tending “food chains.” Occasionally the “downside” is included, but usually only
for lifeforms at the very lowest levels, for instance the “microbial loop” shows a circular
movement pattern for nutrients. Usually the diagrams show all the top organisms as “dead
ends” for nutrients.....perhaps that’s because that’s the way that we unconsciously “want” and “need” to
see them. The fact is that besides feeding off their prey, predators also feed their prey in
more ways than one. It was always the secret to their continued health and success.
But the “productivity” concept gets mixed up with fisheries “production.” A totally
different meaning, “production” here means the amount of flesh extracted from the
ecosystem by humans.
From a text on reef fisheries:
“Because industrial fishing gears are largely precluded, reef fisheries are the domain of
small-scale fishers....Production is governed primarily by two features: distance from
centers of human population, and population densities and hence demand for seafoods.
Many remote Pacific atolls have perhaps never been fished by anything more than a
passing yacht.” (Polunin and Roberts, 1996)
By this definition the Pacific atoll reefs are not particularly “productive,” yet they are
among the healthiest in the world, with the best developed and thriving coral cover, a
feature that should define them as highly “productive.” It is evident that there are two differing
definitions for the word, but it seems that they clearly have the potential to cause a bit of
confusion. One example is in the assumption that since large amounts of fish have been
removed from a reef in the past, that it should always continue to be able to give up large
amounts of fish. “Production” of coral reef fisheries has clearly been declining, sharply in
recent decades but a gradual decline reaching back much farther is also apparent. Has
“primary production” also been declining in these systems? Maybe. It seems possible that it
has been, but reefs tend to only have had single calculations done so no trends have been
revealed. Another question is whether just “primary production”/”carbon fixation by algae” is a
good enough indicator of overall health. It does not appear to be, but that seems to be how it is
interpreted at times. (Reassurances are given that "primary production is as healthy as ever," but they seem rather shaky, and largely based on chlorophyll concentrations.) If “primary production” has gradually declined, and the slowing of
the growth of corals in clean waters seems to suggest that it has...it also seems entirely
possible that it has been an insidious, indirect result of “nutrient” depletion due to
all-fishing.
“Worldwide, the potential sustainable yield of fish, crustaceans, and molluscs from reefs
represents approximately 10% of the world’s annual fisheries take - worth billions of
dollars.” (NOAA)
“...In general, it is clear that degradation has outpaced our comprehension of the problems
at many locations.” (same NOAA source)
Two seemingly contradictory statements - how can the first one really be made with any
confidence?
The conclusions drawn about the coral reef system in a recent marine ecology text:
“Despite all the uncertainties about the relative contributions to the coral-reef ecosystems of
different kinds of autotroph, bacteria, dissolved organic matter, and internal vs. external
inputs, it is clear that the phenomenally high total productivity is in large measure due to
the combination of a tremendous surface area of photosynthetic tissue (either in the form
of zooxanthellae or benthic algae and higher plants), optimal light and temperature
conditions for photosynthesis, and the tight recycling of nutrients in an otherwise
nutrient-poor environment. The efficient recycling of nutrients occurs both at the level of
the coral-zooxanthellae symbiosis and at the general level of the overall food web. Many
consumers are present in reefs and although in absolute terms primary production is high,
relative to the number of consumers food can be regarded as scarce. Hence food is
consumed rapidly and utilization is efficient. A high proportion of the environmental pool
of nutrients is therefore maintained within living tissues, so reducing opportunities for the
loss of nutrients out of the system. Any such losses are compensated by the slow accrual of
nutrients from water masses passing over the reef and by the nitrogen-fixing activities of
blue-green algal associations on the reef or rhizomes of adjacent sea-grasses.” (Barnes and
Hughes, 1999, p 141)
Looking through this for the nutrient balance, it seems that nutrients lost through fishing
must also necessarily be replaced by the “slow accrual of nutrients from water masses
passing over the reef” (water which is well known not to contain a great lot of nutritional
value) and “the nitrogen-fixing activities of blue-green algae.” Therein lies the “hitch,”
since “slow accrual” is all that has ever been managed by natural marine systems
endeavoring to increase their fixed nitrogen/”nutrient” content. “Slow accrual” worked to
build up the rich collection of life on the coral reefs, but “slow” it really was...it took
millions of years. Fishing removals have clearly overwhelmed the ability of these
ecosystems to replace their “nutrient” losses, and now they are impoverished to the point
where corals are dying from starvation in “pristine” water. This occurrence is a “first” in
coral history -- and sadly gives the ominous impression that if some positive change doesn’t
happen soon, it may possibly also be the “last.”
From the introduction to a reef fisheries text:
“... in comparison to lakes, upwellings or ocean shelf systems, the production ecology of
reefs is not well understood. For example, corals entail a subtle symbiosis between zooids
and algae that is hard to quantify. A coral reef has been described as a ‘wall of mouths’
designed to intercept food transported on ocean currents in the midst of low productivity
areas, but we do not clearly understand how to predict growth and production in these
systems. These areas of ecological ignorance mean that the fundamental basis for
assessment, evaluation and management of reef fisheries suffers from serious deficiencies
that impede insight into the effects of harvest.” (Pitcher, T. (ed) in Polunin and Roberts,
1996)
That’s a fairly accurate assessment of the situation. But...the
“understanding” that we think we have about the production ecology of upwelling and
ocean shelf systems, has been based on the idea that these are constantly replenished (and
fishing removals thereby replaced) by a “vast pool of nutrients” in the seawater. No so in
the tropics, there the picture should be much “clearer” - but our inability to really
appreciate the workings of the natural “food cycle” leads us into a major difficulty in
“modelling” these systems and “managing” these fisheries. In fact, the production ecology
models that have been used for upwelling and ocean shelf systems have been falling apart
lately....examples abound, “unexpected” downturns and changes are virtually
everywhere...it seems that the “serious deficiency” is not only in the tropical marine
science. “...insight into the effects of harvest” -- that’s it, that’s the point that needs to be
carefully examined, perhaps from a new angle that has not been previously considered.
And the situation with tropical fish and corals provides a very strong clue as to “what” is
missing. It has been suggested that a coral reef ecosystem might serve as a microcosm for
the oceanic ecosystem as a whole - and that’s not unreasonable. They are as close as
we’ll get to seeing an approximation of a natural “self-contained” marine system, so the
principles that emerge in the coral reef ecosystem are likely to add insight into the workings
of the larger system. Also, the reef system changes should offer insight into the overall
“effects of harvest” in the larger system. And it seems now that the very base of these
systems, the reef building corals, have been damaged.
The corals are bleaching, the fish are fewer, are there any other recent trends in the coral
reef ecosystems? Are there any other parallels with recorded changes in fish stocks
elsewhere?
The stories of the tropical fisheries sound much like those in the rest of the world. Maybe
relatively lacking in recorded data (it’s hard to find “time series” data on anything), but the
themes of a long-term gradual decline to a now very depleted state, declining trophic level
of organisms, declining abundance of fish and average size of fish, and declining size at
sexual maturity...are the same in the tropics as they are wherever we have exploited marine
life. The “disappearance of the big ones” is a major theme all over, and is noted in the reef
fisheries as well. This is said to be because we have preferentially targeted the big ones,
and have thereby been killing them off more quickly than they can replace themselves. But
the explanation may not be as simple as that - for example, there’s contradictory evidence
from the North Atlantic where “big cod” are increasingly vanishing from an unfished stock.
Might another factor, affecting the system overall, be subtly “forcing” the bigger fish to
disappear?
I have only been to the tropics a few times, but I recall often eating fish while I was there.
One thing I remember, that seemed to be common practice, was avoiding the consumption
of the larger specimens of reef fish such as groupers. This is because the bigger ones tend
to accumulate ciguatera - a toxin endemic in tropical fisheries; known for centuries, it is
capable of causing illness in human consumers. I remember being told that the spear
fishermen did not select fish above a certain size for this reason. Eating the big ones just
becomes too risky. I really don’t know whether or not the avoidance of large fish would be
strong enough to have an effect on the population - but it seems that “possibly,” this
practice should have spared a noticeable number of the bigger older fish, and “perhaps”
they should still be there. But they are not, the big groupers are definitely on the “missing
list” for the fished out areas. If systemic food depletion is contributing to the picture
however, the higher trophic level fishes, like the big grouper (and the big cod), will find
themselves increasingly food limited, and this fact alone could account for their
disappearance. Has it? (To compare with the northern cod again, records there show that
the bigger, older fish showed a steeper declining trend in weight-at-age than did the
younger ones...and then the big ones just vanished from the data tables.)
In conclusion, there is at least one more reason why fishing should be considered as a
threat to the wider ecosystem. It seems that fish, even top carnivores, play important roles
in nutrient cycling in coral reef systems; therefore fishing needs to be examined as an
activity that inevitably undermines and decreases nutrient availability overall (...and concepts of “nutrient” need to move beyond the liquid forms).
Therefore, the next section of this report examines the pathophysiology of coral bleaching, pursuing a hunch that
the problem might stem from a shortage of “nutrients.”

 The secret to the success of the coral reefs is commonly believed to be the “tight
recycling of nutrients” in the system, particularly in the corals, in which tiny plants and
animals live together in a symbiosis that conserves key nutrients quite effectively. Algae
(zooxanthellae), living inside the tissues of the cnidarian host, harness energy from sunlight
and “fix carbon” by photosynthesis. Energy from this source is provided to the polyp host
in return for exclusive access to the waste-nutrients produced by the host. These wastes (N
and P) function to fertilize the algae. The most significant, and apparently “limiting”
nutrient in the picture is fixed nitrogen, a critical element in the construction of all proteins.
The symbiotic arrangement allows the partners to avoid the loss of fixed N to the water, a
process that normally occurs in both free floating algae and solo-living marine animal
lifeforms. In larger marine systems, overall fixed N is essentially passed back and forth
between the plant and animal compartments of the community - corals have evolved a way
to capture and complete this loop inside their own special symbiosis. It’s a micro-model of
what happens in the bigger picture, and the same principle applies in the more “nutrient”
rich systems of the temperate and polar zones. Plants and animals perpetually passing the
precious ball (fixed N) back and forth - it’s a common theme in all living systems on the
planet.
The secret to the success of the coral reefs is commonly believed to be the “tight
recycling of nutrients” in the system, particularly in the corals, in which tiny plants and
animals live together in a symbiosis that conserves key nutrients quite effectively. Algae
(zooxanthellae), living inside the tissues of the cnidarian host, harness energy from sunlight
and “fix carbon” by photosynthesis. Energy from this source is provided to the polyp host
in return for exclusive access to the waste-nutrients produced by the host. These wastes (N
and P) function to fertilize the algae. The most significant, and apparently “limiting”
nutrient in the picture is fixed nitrogen, a critical element in the construction of all proteins.
The symbiotic arrangement allows the partners to avoid the loss of fixed N to the water, a
process that normally occurs in both free floating algae and solo-living marine animal
lifeforms. In larger marine systems, overall fixed N is essentially passed back and forth
between the plant and animal compartments of the community - corals have evolved a way
to capture and complete this loop inside their own special symbiosis. It’s a micro-model of
what happens in the bigger picture, and the same principle applies in the more “nutrient”
rich systems of the temperate and polar zones. Plants and animals perpetually passing the
precious ball (fixed N) back and forth - it’s a common theme in all living systems on the
planet.
 There it is again, “the top of the food chain”...as if there is nowhere to go from there,
except possibly “up” another notch to be consumed by humans (?). And what exactly is the
meaning to be taken from the word “productivity” in that sentence? There’s the myth
again, that the creatures at the higher trophic levels are not particularly important to the
functioning of the ecosystem as a whole. But the truth is that they were simply living there
and practicing “efficient recycling of nutrients,” just like all their neighbours at the “lower trophic levels.”
There it is again, “the top of the food chain”...as if there is nowhere to go from there,
except possibly “up” another notch to be consumed by humans (?). And what exactly is the
meaning to be taken from the word “productivity” in that sentence? There’s the myth
again, that the creatures at the higher trophic levels are not particularly important to the
functioning of the ecosystem as a whole. But the truth is that they were simply living there
and practicing “efficient recycling of nutrients,” just like all their neighbours at the “lower trophic levels.”